ТэаЁефЃЈвыЃЉ бІЯўЩњ ЭѕБЬСЋ жм ЮЇ вІалОќЃЈаЃЃЉ
еЊ вЊ аЁыФКЭгЮРыАБЛљЫсЪЧШеСИЕААздкаЁГІжажївЊЕФЯћЛЏВњЮяЃЌФПЧАЩаВЛУїШЗетСНжжЮяжЪМфЕФзЊдЫИЩШХЁЃБОЪдбщгУCaco-2ЯИАћРДбаОПPEPT1зЊдЫЕФЖўыФЮќЪеЪЧЗёЛсИФБфгЮРыЫсадЁЂжаадАБЛљЫсЕФЮќЪеЁЃL-[3H]ArgзЊдЫЕНCaco-2ЯИАћЪЧЗЧвРРЕNa+ЃЌЖјжївЊЪЧЪмb0ЃЌ+зЊдЫЯЕЭГЕФЕїНкЁЃ10 mmol/lЖўыФХрбјЕФЯИАћФмНЋL-ArgСїЫйЬсИпЕН4БЖЁЃБОЪдбщЪзДЮжЄУїЃЌаЁГІЯИАћФкгЮРыАБЛљЫсКЭаЁыФЕФЮќЪеЯрЛЅгАЯьЃЌВЂФмИФБфАБЛљЫсЮќЪеЕФЖЏСІбЇЁЃ
ЙиМќДЪ ЖўыФPEPT1ЃЛboЃЌ+ЯЕЭГЃЛГІЕРАБЛљЫс
жаЭМЗжРрКХ Q517
ШеСИЕААзКЭФкдДадЕААзОвШЕААзУИЁЂаЁГІЫЂзДдЕЕААзУИКЭыФУИЯћЛЏКѓаЮГЩаЁыФКЭгЮРыАБЛљЫсЁЃаЁГІЯИАћЖЅЖЫЯИАћФЄФкМИжжжБНгЕФзЊдЫЯЕЭГЕїНкгЮРыАБЛљЫсЕФЮќЪеЃЌЕЋЖўыФгыШ§ыФЕФЮќЪеЪЧЛљгкPEPT1ЯЕЭГЭъГЩЁЃЫфШЛыФгыАБЛљЫсЕФзЊдЫОљЛсЕМжТАБЬЌЕЊЮќЪеШыаЁГІЃЌЕЋВЛЭЌзЊдЫЯЕЭГЖдАБЛљЫсЮќЪеЕФгАЯьШдВЛУїШЗЁЃЫфШЛАБЛљЫсВЛЛсгАЯьPEPT1ЯЕЭГзЊдЫыФЃЌЕЋЩаВЛУїШЗЯИАћЫЎНтКѓыФЕФЮќЪеЪЧЗёЛсгАЯьгЮРыАБЛљЫсНјШыЩЯЦЄЯИАћЁЃCaco-2ЯИАћРДдДгкШЫЬхГІЕРЩЯЦЄЯИАћЃЌЪЧвЛИігаМлжЕЕФЬхЭтбаОПФЃаЭЃЌПЩвдФЃФтЫсадЁЂМюадЁЂжаадАБЛљЫсзЊдЫЯЕЭГМАPEPT1зЊдЫЯЕЭГЁЃ
БОДЮбаОПжаЃЌЮЊСЫФмФЃФтВЛЭЌЕФзЊдЫЯЕЭГЃЌЪдбщВЩгУЕФЪЧШкКЯКѓ14 dЕФCaco-2ЯИАћЁЃЪзЯШЃЌЪдбщУїШЗСЫВЛЭЌзЊдЫЯЕЭГЖдЗХЩфадБъМЧЕФЫсадзѓа§ОЋАБЫс(L-Arg)ЁЂжаадзѓа§БћАБЫс(L-Ala)ЮќЪеЕФгАЯьЁЃБОДЮЪдбщжївЊФПЕФЪЧбаОПЯИАћОгУКЌгаОЋАБЫсЁЂБћАБЫсЕФЖўыФДІРэКѓЃЌЖдL-Argгы L-AlaзЊдЫТЪЕФгАЯьЁЃЮЊСЫБугкбаОПЖўыФЖдАБЛљЫсСїЫйЕФгАЯьЃЌЪдбщгІгУНКдЕААзХрбјЕФCaco-2ЯИАћЃЌНЕЕЭЗЧЬивьадзЊдЫЯЕЭГЖдАБЛљЫсЁЂыФзЊдЫЕФгАЯьЁЃ
1 ВФСЯКЭЗНЗЈ
1.1 ВФСЯ
КЯГЩD-[3H]Phe-L-Ala(9 Ci/mmol)КЭЮДБъМЧЕФD-Phe-L-AlaЗжБ№ЙКздBiotrendКЭBachemЙЋЫОЃЌL-[3H]ArgЃЈ50 Ci/mmolЃЉгЩAmershamЙЋЫОЬсЙЉЃЌL-[3H]Ala (96 Ci/mmol)гЩICNЬсЙЉЃЌЦфЫќЕФАБЛљЫсКЭыФОљЙКздSigmaЙЋЫОЁЃЯИАћХрбјУѓКЭХрбјЦПЙКздRennerЙЋЫОЃЌЯИАћХрбјЫљашЕФЦфЫќВФСЯРДздGibcoЙЋЫОЃЌНКдЕААзЙКздServaЙЋЫОЁЃ
1.2 ЯИАћХрбј
Caco-2ЯИАћ(ATCC)ХрбјЁЂДЋДњОљгУDMEMХрбјвК (КЌ10%ЬЅХЃбЊЧхЁЂ2 mmol/lЙШАБѕЃАЗЁЂ1% MEMЗЧБиашАБЛљЫсЁЂ70 ІЬg/mlЧьДѓУЙЫи)ЃЌдк37 ЁцЁЂ5% CO2ЬѕМўЯТНјааЁЃ
1.3 зЊдЫбаОП
Caco-2ЯИАћЕФСїЖЏбаОПЪЧдкKrebsЛКГхЦїжаНјааЃЌЦфФкХфга137 mmol/l NaClЁЂ5.4 mmol/l KClЁЂ2.8 mmol/l CaCl2ЁЂ1.0 mmol/l MgSO4ЁЂ0.3 mmol/l NaH2PO4ЁЂ0.3 mmol/l KH2PO4ЁЂ10 mmol/lЦЯЬбЬЧКЭ10 mmol/l HEPES/Tris(pHжЕ7.4)ЁЃдкЮоNa+ЛКГхЦїжаЃЌ137 mmol/lТШЛЏЕЈМюЬцДњ137 mmol/l NaClЃЌ0.3 mmol/l KH2PO4ЬцДњ0.3 mmol/l NaH2PO4ЃЌЦфЫќГЩЗжЯрЭЌЁЃбаОПЮќЪеТЪЪБЃЌЯДЭбХрбјдк24ПзАхжаЕФЕЅВуЯИАћвдШЅГ§ХрбјвКЃЌШЛКѓдк37 ЁцЯТгыОКељЕзЮяКЭЗХЩфадБъМЧЕзЮяЛђНігыЗХЩфадБъМЧЕзЮявЛЦ№Хрбј30 min,ШЛКѓгУБљЕуЕФХрбјЛКГхвКЯДЭб3ДЮ,УППзМг50 ІЬl TENЛКГхвКЃЌВЂМг50 ІЬlзщжЏдіШмМСНјааЯћЛЏЁЃЭЈЙ§МьВтЯИАћЗХЩфадРДШЗЖЈЪОзйдЊЫиЕФЕМШыЁЃЯИАћХрбјЬѕМўЃКХрбјЪБМфЮЊ30 minЃЌХрбјЮТЖШ37 ЁцЃЌЖўыФМАЖўыФзщГЩАБЛљЫсЕФХЈЖШОљЮЊ10 mmol/lЁЃ
1.4 ЭГМЦ
ЫљгаЗжЮі(ЯпадЛђЗЧЯпадЛиЙщЗжЮі)ОљгУPrism2.01ЭъГЩЃЌЪ§ОнвдЦНОљжЕЁРБъзМЮѓ(SEM)БэЪОЃЌВювьЯджјадгУStudent tМьбщЁЃ
2 НсЙћгыЗжЮі
2.1 Caco-2ЯИАћжаL-[3H]ArgКЭL-[3H]AlaЕФзЊдЫЬиЕуЃЈМћЭМ1ЁЂЭМ2ЃЉ
A
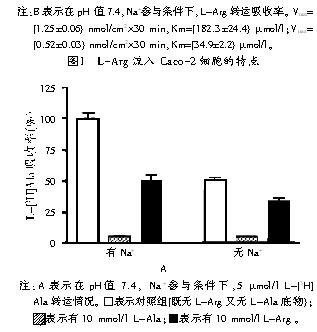

АБЛљЫсзЊдЫЪдбщЪЧдкpHжЕ7.4ЬѕМўЯТНјааЕФЃЌвдНЕЕЭH+вРРЕзЊдЫЬхЯЕЖдАБЛљЫсЮќЪеЕФгАЯьЁЃЮоТлЪЧЗёгаNa+ЕФВЮгыЃЌдк10 mmol/l L-ArgЪдбщзщжаЃЌ5 ІЬmol/l гаЗХЩфадБъМЧЕФЫсад L-[3H]ArgЮќЪеТЪЖМЯТНЕЕНЖдеезщЕФ7% (МћЭМ1A)ЁЃдкNa+ВЮгыЯТЃЌ10 mmol/l L-AlaЪдбщзщгыЖдеезщЯрБШЃЌ5 ІЬmol/l гаЗХЩфБъМЧЕФЫсад L-[3H]ArgЮќЪеТЪЯТНЕСЫ83%(МћЭМ1A)ЃЛдкМШЮоL-ArgгжЮоL-AlaЕзЮяЕФЖдеезщжаЃЌдкЮоNa+ВЮгыЪБ5 ІЬmol/l L-[3H]ArgЕФЮќЪеТЪЪЧгаNa+ЕФЬѕМўЯТЕФ2БЖ(МћЭМ1A)ЁЃЮоNa+ВЮгыЯТЃЌЫсадАБЛљЫсЃЈШчL-LysЃЉЮќЪеТЪИќИпЃЌетЪЧгЩгкЕчУєИаЛњжЦв§Ц№ЕФЃЌИУЛњжЦЛсЪмЕНNa+ЕФвжжЦЁЃL-Ala(10 mmol/l)НЕЕЭСЫЗЧвРРЕNa+ L-[3H]Arg ЮќЪеТЪЕФ85%ЃЌетБэУїАБЛљЫсЕФвЦЖЏЪЧгЩb0ЃЌ+ЯЕЭГзЊдЫЕФЃЌИУЯЕЭГПЩдкЮоNa+ЕФЬѕМўЯТзЊдЫЫсадЁЂжаадАБЛљЫсЁЃ15%ЕФL-[3H]ArgЮќЪеЪЧВЛЪмL-AlaЕФгАЯьЃЌЖјЪЧгЩY+ЯЕЭГ(ВЛвРРЕNa+ПЩзЊдЫЫсадАБЛљЫс)зЊдЫЛђепЪЧгЩY+LЯЕЭГ(дкЮоNa+ЕФЬѕМўЯТзЊдЫЫсадАБЛљЫсЃЌдкгаNa+ЕФЬѕМўЯТзЊдЫжаадАБЛљЫс)зЊдЫЁЃ
ЮЊЦРЖЈL-ArgЕФЮќЪеЪЧЗёБЛетСНжжзЊдЫЭЈЕРЫљЕїНкЃЌЮвУЧзїГіСЫвЛИіСїЫйгыЕзЮяХЈЖШМфЕФКЏЪ§ЭМЁЃзЊдЫТЪБэЪОБЅКЭЖЏСІбЇЃЈМћЭМ1BЃЉЃЌЫйТЪЕФБфЛЏБэЪОЯрЙиадЕФИпЕЭЃЌетСНжжзЊдЫЭЈЕРЕФKmжЕЗжБ№ЪЧЃЈ182.3ЁР24.4ЃЉ ІЬmol/lЁЂЃЈ34.9ЁР2.2ЃЉ ІЬmol/lЃЈМћЭМ1BЃЉЁЃ
L-[3H]AlaзЊдЫЕНCaco-2ЯИАћжївЊЪмзЊдЫЯЕЭГЕФЕїНкЖјВЛЪмL-[3H]ArgЮќЪеЕФгАЯьЁЃЮоNa+ВЮгыЯТЃЌL-[3H]AlaЕФзЊдЫНіЪЧгаNa+ЬѕМўЯТЕФвЛАы(МћЭМ2A)ЁЃЮоNa+ЬѕМўЯТЃЌ10 mmol/l L-ArgвжжЦСЫ30%ЕФL-[3H]AlaЮќЪе(МћЭМ2A)ЁЃдкгаNa+ЬѕМўЯТЃЌL-[3H]AlaЕФЮќЪеЪЧЮоNa+ЬѕМўЯТЕФ2БЖЃЌетЪЧгЩгкNa+ДцдкЪБЃЌb0ЃЌ+ЯЕЭГзЊдЫСЫ15% L-AlaЁЃбаОПБэУїЃЌМИжжзЊдЫЯЕЭГОљВЮгыСЫL-AlaЕФЮќЪезЊдЫЃЌL-[3H]AlaЫйТЪЖЏСІбЇЯдЪОKmжЕЛљБОЪЧвЛГЃЪ§ЃЌдМЮЊ150 ІЬmol/l(МћЭМ2B)ЃЌетвВЫЕУїВЮгы L-AlaЮќЪезЊдЫЕФВЛЭЌзЊдЫЯЕЭГЕФKmжЕЛљБОЯрЭЌЁЃ
2.2 ЖўыФКЭгЮРыАБЛљЫсДІРэЙ§ЕФCaco-2ЯИАћЖдL-[3H]AlaЁЂL-[3H]ArgСїЫйЕФгАЯь
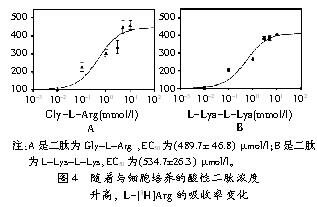
ЖўыФХЈЖШВЛЖЯЩ§ИпЕФЧщПіЯТЃЌВтСП5 ІЬmol/l DЃ[3H]Phe-L-AlaЮќЪеТЪЃЌДгЖјбаОПЖўыФгыPEPT1ЕФЯрЙиад(МћЭМ3)ЁЃEC50жЕЗжБ№ЪЧЃКL-Lys-L-LysЮЊЃЈ411.1ЁР15.3ЃЉ ІЬmol/lЃЌGly-L-ArgЮЊЃЈ412.7ЁР13.7ЃЉ ІЬmol/lЃЌGly-L-HisЮЊ(42.0ЁР6.5) ІЬmol/lЃЌL-Ala-L-AlaЮЊ(271.3ЁР12.4) ІЬmol/l(МћЭМ3)ЁЃЩЯЪіШЮвЛжжЖўыФХЈЖШЮЊ10 mmol/lЪБЃЌзЊдЫЯЕЭГЕФзЊдЫЫйЖШОљДязюДѓжЕVmaxЁЃдкЧАДІРэЙ§ГЬжаЃЌЗжБ№гУ10 mmol/l L-Lys-L-LysЁЂGly-L-ArgЁЂGly-L-His ЁЂL-Ala-L-Ala ХрбјCaco-2ЯИАћЃЛЭЌЪБгУЭЌФІЖћХЈЖШЕФАБЛљЫс(ЖўыФЕФзщГЩАБЛљЫс)НјаадЄХрбјЃЌВЂБШНЯЖўыФгыгЮРыАБЛљЫсзЊдЫЯЕЭГМфЕФгАЯьзїгУЁЃвѕадЖдее(ЕзЮяЮЊ10 mmol/l Gly-GlyЛђ20 mmol/l Gly)ЫЕУїb0ЃЌ+ЯЕЭГЖдL-[3H]ArgЮќЪеЕФгАЯьЃЌШЛКѓМЦЫуL-[3H]Arg(50 Ci/mmol)КЭL-[3H]AlaНјШыCaco-2ЯИАћЕФЫйЖШЁЃ

Г§СЫGly(ЮоТлЪЧвдЖўыФаЮЪНЛЙЪЧвдгЮРыаЮЪН)ЭтЃЌЦфЫќШЮвЛжжЧАХрбјЗНЪНОљУїЯдЬсИпСЫCaco-2ЯИАћЖдL-[3H]ArgЕФЮќЪеЫйЖШ(МћБэ1)ЁЃгУЖўыФХрбјКѓЕФЮќЪеЫйЖШНЯАБЛљЫсЕФПьЃЌЖўыФгыАБЛљЫсЛьКЯЮяХрбјКѓЮќЪеЫйЖШзюПьЁЃгУGly-L-ArgЁЂGlyЁЂL-ArgЛьКЯЮяЛђL-Ala-L-AlaЁЂL-AlaЛьКЯЮяЧАХрбјCaco-2ЯИАћКѓЃЌL-[3H]ArgЕФЮќЪеЫйЖШЬсИпСЫ4.6БЖ(МћБэ1)ЁЃгыL-[3H]ArgзЊдЫЯрЗДЃЌгУL-Ala-L-AlaЁЂGly-L-ArgЁЂL-Lys-L-Lys ЛђЦфзщГЩАБЛљЫсЛђЖўыФгыАБЛљЫсЕФЛьКЯЮяХрбјCaco-2ЯИАћКѓЃЌL-[3H]AlaЕФЮќЪеЫйЖШОљУЛгаЯджјЬсИп(МћБэ2)ЁЃ


2.3 ЖўыФАћФкЫЎНтДйНјL-[3H]ArgЮќЪе
вдЯТЪдбщжЄУїСЫЖўыФАћФкЫЎНтЪЧL-[3H]AlaЕФЮќЪеЧАЬсЬѕМўЃКЂйЗЧЫЎНтЕФD-Phe-L-Ala ВЛФмДйНјL-[3H]AlaЕФзЊдЫ(МћБэ3)ЃЛЂкЯШгУАБыФУИвжжЦМС(Amastatin)ЧАДІРэЯИАћвдвжжЦАћФкЫЎНтЃЌШЛКѓдйгыGly-L-ArgНјааЧАХрбјвжжЦАћЭтЫЎНтКѓЃЌВЛФмДйНјзЊдЫ(МћБэ3)ЁЃAmastatinЙуЗКгУзїыФЫЎНтвжжЦМСЃЌЧвЦфВЛЛсИЩШХЩЯЦЄЯИАћыФзЊдЫЯЕЭГЃЌвђДЫAmastatinЪЧЭЈЙ§вжжЦGly-L-ArgЕФЫЎНтзшАGly-L-ArgЕФЮќЪеЁЃЯИАћОгЮРыАБЛљЫсЧАДІРэКѓЃЌAmastatinВЛФмгАЯьЯИАћПчФЄзЊдЫЃЌетвВЫЕУїСЫAmastatinЖдЫЎНтЕФЬиЪтзїгУ(МћБэ3)ЁЃ

L-[3H]ArgЮќЪеЕФЯоЫйВНжшЪЧЖўыФзЊдЫЕНЯИАћЛЙЪЧЖўыФАћФкЫЎНтФиЃПЪдбщжЄУїЃЌЯоЫйВНжшЪЧзЊдЫЖјВЛЪЧЫЎНт(МћЭМ4)ЁЃЭЈЙ§Gly-L-ArgЛђL-Lys-L-LysЧАДІРэЯИАћЪдбщЗЂЯжL-ArgСїЫйДя1/2 VmaxЪБGly-L-ArgЪЧ(489.7ЁР46.8) ІЬmol/lЃЌL-Lys-L-LysЪЧ(534.7ЁР26.3) ІЬmol/l (МћЭМ4)ЁЃL-[3H]Arg ПчФЄзЊдЫEC50жЕЕФДѓаЁЯрЫЦгкЭЌPEPT1зїгУЕФыФЕФв§СІ(МћЭМ3)ЁЃ
3 ЬжТл
аЁГІжаАБЬЌЕЊЮќЪеМШвдгЮРыаЮЪНЃЌгжвдЖўыФЁЂШ§ыФЕФаЮЪНЃЌЕЋВЛЭЌЕФзЊдЫЯЕЭГЖдАБЛљЫсЮќЪегАЯьГЬЖШЩаВЛУїШЗЁЃвдЭљЪдбщУЛгадкЯИАћЫЎЦНбаОПАБЛљЫсКЭЖўыФЮќЪеЕФЯрЛЅзїгУЃЌвђетжжбаОПЪЪвЫгкдкРрЫЦЬсШЁЯИАћЕФзщжЏЛЗОГЯТНјааЁЃБОЪдбщбЁгУЕФЪЧCaco-2ЯИАћЁЃ
ЭЈЙ§Caco-2ЯИАћбаОПЪдбщЃЌжЄУїСЫЖўыФзЊдЫКЭЫсадАБЛљЫсзЊдЫМфЕФЯрЛЅзїгУЁЃЫсадL-LysЪмЕНСНжжзЊдЫЯЕЭГЕФЕїНкЃЌвЛИіЪЧЗЧвРРЕNa+ЫсадЁЂжаадАБЛљЫсзЊдЫЯЕЭГЃЌСэвЛИіЪЧЬиЖЈАБЛљЫсзЊдЫЯЕЭГЁЃдквдЧАбаОПжаЃЌЕквЛИіЯЕЭГМДb0ЃЌ+ЯЕЭГФмзЊдЫ47%L-LysЃЌЬиЖЈЫсадАБЛљЫсзЊдЫЯЕЭГY+зЊдЫ27%ЃЌХЈЖШВюзЊдЫ26%ЁЃ64%ЪЧгЩдиЬхзЊдЫЯЕЭГb0ЃЌ+зЊдЫЃЌ36%гЩЯЕЭГY+зЊдЫЁЃдкБОДЮбаОПжаЃЌL-[3H]Arg 85%гЩЯЕЭГb0ЃЌ+ЕїНкЃЌ15%гЩЯЕЭГY+ЕїНкЁЃОЯЕЭГb0ЃЌ+зЊдЫL-ArgЕФБШТЪИќИпЃЌдвђЪЧЯИАћВЛЭЌЁЂЩњГЄвђзгВЛЭЌЁЃ
ЫсадЁЂжаадАБЛљЫсЮќЪеБиаывЊгаЯЕЭГb0ЃЌ+зЊдЫЃЌЖдL-AlaЮќЪеБШвРРЕNa+зЊдЫЕЭ15%ЁЃдкNa+ДцдкЯТЃЌзЊдЫжаадЁЂЫсадАБЛљЫсЕФb0ЃЌ+ЯЕЭГжЛзЊдЫL-Ala ЮќЪезмСПЕФ35%ЁЃb0ЃЌ+зЊдЫЯЕЭГШдЮДдкГІЕРзщжЏЗжзгЫЎЦНЩЯЕУвджЄЪЕЁЃдУћЮЊNBBзЊдЫЯЕЭГИќУћЮЊЯЕЭГbЃЌФмИќКУЕиЗДгГГігыЯЕЭГb0ЃЌ+ЕФЙиЯЕЁЃ
ЯдШЛЃЌгыL-ArgМАЦфзЊдЫЯЕЭГЯрЗДЃЌL-AlaЕФСїЫйВЛЪмЧАХрбјЮя(ЮоТлЪЧЫсадЁЂжаадАБЛљЫсЛЙЪЧЖўыФ)ЕФгАЯьЁЃ15%L-AlaЮќЪеСПЪЧЯЕЭГb0ЃЌ+зЊдЫЃЌФкдДАБЛљЫсВЛФмЯджјгАЯьL-AlaСїЫйЁЃ
БОДЮЪдбщЕквЛДЮдкЯИАћЫЎЦНжЄЪЕаЁыФгыгЮРыАБЛљЫсЮќЪежЎМфЯрЛЅзїгУЃЌВЂЮЊPEPT1ЖдАБЛљЫсЮќЪеЕФЕїНкзїгУЬсЙЉСЫжЄОнЁЃаЁыФзЊдЫЯЕЭГПЩзЊдЫ400ЖржжЖўыФКЭ8 000жжШ§ыФЃЌФмЯджјИФЩЦГІЕРгЮРыАБЛљЫсЕФзЊдЫЁЃPEPT1ПЩзЊдЫДѓСПаЁыФЕНГІЕРЩЯЦЄЯИАћЃЌетаЉаЁыФЫЎНтКѓЪЭЗХГігЮРыАБЛљЫсЃЌДгЖјДйНјБиашАБЛљЫсШчL-ArgЁЂL-LysЕФЮќЪеЁЃаЁыФзЊдЫЯЕЭГФмДйНјГІЕРАБЛљЫсЕФЮќЪеЁЃЪзЯШЃЌЖўыФЁЂШ§ыФ(КЌгаБиашАБЛљЫс)ЕФзЊдЫБмУтСЫБиашАБЛљЫсВЮгыАБЛљЫсзЊдЫЯЕЭГЕФОКељЃЛЦфДЮЃЌгЩгкдіМгЯИАћФкгЮРыАБЛљЫсКЌСПЃЌДгЖјФмДйНјБиашАБЛљЫсЕФЮќЪеЁЃ
ЃЈБрМЃКСѕУєдОЃЌЃЉ
ТэаЁефЃЌИЃНЈГіШыОГМьбщМьвпОжЃЌ850001,ИЃжнЪаКўЖЋТЗ312КХЁЃ
бІЯўЩњЁЂЭѕБЬСЋЁЂжмЮЇЁЂвІалОќЃЌЛЊДяЩњЛЏПЦММЫЧСЯ(КўББ)гаЯоЙЋЫОЁЃ
ЪеИхШеЦкЃК2007-01-23
зЂЃКвыздЯИАћЩњРэбЇдгжОЃЌ2001ЃЈ186ЃЉЃК251ЁЋ259 |














